|
FECAL DNA:
STUDIES USING NON INVASIVE TECHNIQUES IN POPULATIONS OF WILD PRIMATES IN LABORATORIO DE PRIMATOLOGIA Estación de Biología "Los Tuxtlas" Instituto de Biología-UNAM |
|
|
|

Genetic studies through fecal DNA
Introduction
Population geneticists are relying
increasingly on the study of DNA variation within and among populations (5). In
the past, traditional genetics were dependent on visible differences between
species. Usually only domesticated animals or cultivated plants could be used
because controlled crosses had to be done to be able to do analyses by using Mendel´s principles of inheritance (8). With the coming of
molecular techniques, genetic differences can now be obtained by direct
analyses of DNA and proteins. The use of DNA in population genetics gives an
enormous amount of possibilities to answer research questions. Genetic variation within and between
populations, divergence between species and questions
on phylogeny can be answered. Besides this, DNA fingerprinting can be done by
which individual animals can be traced to answer questions on home range,
territory and patterns of migration. Pedigree studies can be performed and
histories of populations can be examined (8).
Non-invasive
monitoring
New techniques have been developed to add
to the possibilities to study primates. One of them is the use of non-invasive
monitoring. With this technique DNA of mammals can be collected in the field
without disturbing them. This can be done by collecting faeces
of which DNA can be extracted. DNA is present in faeces
because the intestinal lining is renewed every week, which causes body-cells to
be excreted with the faeces. The extracted DNA can be
analyzed using microsatellite-analysis. This
technique enables researchers to individually type samples, although
observations still have to be done to determine the origin of the sample.
Besides the information DNA provides, other elements in faeces
like hormones or parasites can be examined. The diagram below shows an overview
of the possibilities.
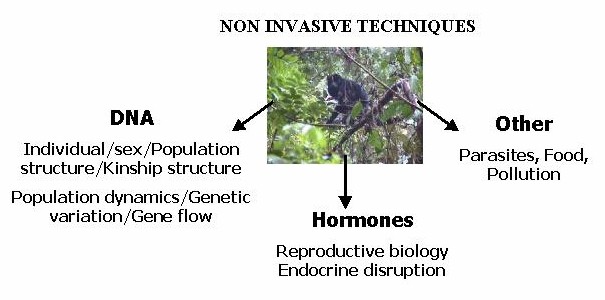
Although
the technique can be very useful, there are some disadvantages, especially when
collecting fecal material. The most important is the small amount of DNA
present in the feces. The DNA can be highly degraded due to passing through the
intestinal track.The above indicates that with DNA,
genetic studies can be done on any organism thinkable. However, when doing
research on animal populations, blood or tissue samples for DNA can be hard to
obtain. Capturing often causes a considerable amount of stress. Nocturnal
animals, marine mammals or animals living in dense forest are particularly
difficult to sample because they are hard to trace (4). Noninvasive sampling
provides solutions for these problems. With this sampling method, DNA material
can be obtained without capturing the animals. DNA is present in the roots of
hairs, in feathers and in feces. Fecal material contains DNA because the
epithelial cells of the intestinal lining are renewed regularly. The old
epithelial cells are excreted with the feces. By collecting these materials,
DNA can be extracted and analyzed. For herbivorous animals, the secondary compounds
present in plants can have an inhibitory effect on the quality of the DNA. The
table below gives an overview of the advantages and disadvantages of using
noninvasive techniques
Table
1: an overview of the advantages and disadvantages of noninvasive sampling
Material
|
Advantages |
Disadvantages |
|
Hairs/feathers |
Provides very useful genetic information |
Hairs are difficult to get (2) |
|
|
Capture not (always) necessary (6) |
|
|
|
Genetic studies difficult due to arboreal life many animals (2) |
|
|
Feces |
Provides very useful genetic information |
Limited amount of DNA (3) |
|
|
Capture not (always) necessary (6) |
Lack of microsatellite primers (2) |
|
|
Genetic studies difficult due to arboreal life many animals (2) |
Degradation of DNA (3) |
|
|
|
Inhibiting secondary compounds |
|
|
The use of non-invasive techniques to
extract DNA has allowed primatologists to recently
carry out studies of genetic variation, gene flow, and paternity, among
other, in populations of primates in the wild. The collecting of fecal DNA
from epithelial cells and the characterization of genetic markers (microsatellites) allow the identification of individuals,
of their sex and of their contribution to the genetic pool in the population.
Thus, these techniques have resulted in evidence about individual
reproductive choices, about kinship structure and about population genetics
(genetic variation and gene flow) in groups and populations of primates. |

|
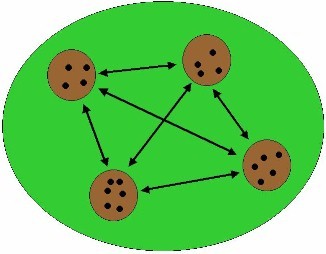
CONTINUOUS FOREST
|
In undisturbed areas, migration can occur between
populations of a specific species. Individuals can migrate to another
population and reproduce. In this way, exchange of genes is possible. This
maintenance of genetic variation is important and beneficial to a population.
When environmental circumstances change, a population should answer to this
by adjusting itself. Natural selection of the fittest individuals will occur.
When genetic variation is high enough, the right combination of genes will be
present in the population. Individuals with these genes will reproduce
offspring, and the population will remain. The diagram to the left
illustrates the "normal" process of gene flow in a population of
primates living in extensive forest tracts. Individuals, males and females,
leave their natal troop and join other troops where they reproduce. When the
forest is fragmented, primate populations are also fragmented and the spatial
separation among forest patches may disrupt the processess
of emigration and immigration among remaining troops. Reduction of genetic
variation, in animal generations, as a result of lack of "fresh"
genes and inbreeding is the outcome. |
|
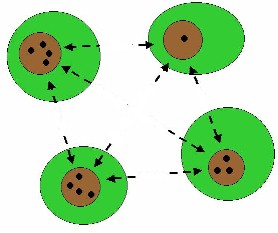
FRAGMENTED FOREST
|
Human destruction of forests causes fragmentation. Because
of this, populations of animals or plants from the same species can become
isolated from each other, thus reducing the possibilities for migration and
genetic transfer. Organisms can become separated from resources or required
habitats by fragmentation, thus reducing individual fitness. More important,
habitat fragmentation is likely to increase extinction proneness due to
inbreeding because small, isolated populations will become inbred overtime (Frankham, 1998). Because of inbreeding, populations will
be less capable of adjusting to new situations. Inbreeding also increases the
occurrence of diseases: dangerous genes are rare in populations with high
genetic variation, but can take over when variation is low. Fragmented areas
are also endangered by edge-effects. Because of the more open area, predation
and habitat deterioration at the edges may be more intense. |
The focus of our genetic research with with
populations of wild primates in southern Mexico is to investigate, by using
non-invasive techniques, the genetic variation and gene flow in populations of
howler and spider monkeys existing in fragmented habitats and in extensive
forest tracts. Forest fragmentation by human activity has resulted not only in
significant reductions in the size of primate populations, but also in
alterations in their demographic structure. As a corollary to these changes,
physical and temporal isolation has diminished the flow, through processes of
emigration and immigration, of individuals among remaining social units, a
situation that may also contribute to a higher incidence of inbreeding. In the
case of populations existing in extensive and protected forest tracts, the
primateâs sociobiological events that lead to
the emigration and immigration of males and females among troops in the
population sustain gene flow and thus adequate genetic variation.
|
|
Our specific interest is to assess, through DNA
extraction from primate fecal material, genetic variation and gene flow in
primate populations existing in extensive forest tracts and in fragmented
landscapes in particular regions of southern |

|
Evidence from
well-designed studies supports the utility of corridors as a conservation
tool (Beier & Noss,
1998). By connecting the isolated populations, migration can occur between
them. Genetic transfer is again possible which increases fitness of the
populations. Edge effects may be reduced by allowing vegetation to grow along
the forest edges to increase cover (Estrada et al. 2002). |
|
|
|
|
DNA extraction is done from the epithelial cells
lining the intestine, cells which are carried by the fecal material as it is
expelled from the digestive tract. About |
|


Field techniques for collecting feces
At the Biological station at Los Tuxtlas and in other sites in southern
Table
2: an overview of the advantages and disadvantages of field techniques for
collecting fecal material
|
Method |
Advantages |
Disadvantages |
|
Use fresh samples |
Higher success rate due to less degradation |
More difficult to find |
|
Process in field or in lab? |
Higher success rate. Less degradation when processed in field |
More difficult to bring gear to the field |
|
Use outer part of feces (1) |
More epithelial cells (1) |
Not possible when feces is scattered |
|
Work sterile (gloves, clean with alcohol) |
Less contamination |
|
|
Process as soon as possible (1) |
Less degradation |
|
Storage techniques for feces
Storing
the samples for further analyses should be done very carefully. Degradation of
DNA can occur easily. Degrading compounds in the feces have to be eliminated in
order to conserve the DNA. There is a variety of techniques used to store fecal
material. Below several techniques are listed (table 3).
Table
3: an overview of the advantages and disadvantages of storage techniques for
fecal material
|
Method |
Advantages |
Disadvantages |
|
95% Ethanol |
Most commonly used (5), cheap, good results (5) |
|
|
Silica gel |
High success rate |
Expensive |
|
Humidifier |
Cheaper than silica gel |
|
|
Freeze-drying |
|
Labor intensive |
|
Buffers from special kits (eg. Qiagen) |
Especially developed for fecal material, high success rate |
Expensive |
|
20% dimethylsulfoxide, saturated with NaCl (5) |
Storage at ambient temperature (5) |
|
Processing
To be able to do analyses, the samples
have to be processed. First, the DNA present in the fecal samples has to be
isolated from the fecal material. Standard techniques are available for this
(5). Because of the small amounts of DNA in the feces, amplification of the DNA
is necessary to obtain larger amounts of DNA. With the Polymerase Chain
Reaction (PCR) DNA can be amplified. Often amplification of microsatellites
is done. These are small pieces of DNA. They are highly variable, so they can
be used for individual typing. More explanation on microsatellites
can be found below.
Microsatellite analysis. Microsatellite
loci (also called Simple Sequence Repeats (SSRs)) are
stretches of nuclear DNA, which are composed of tandemly
repeated units of 2 to 6 base pairs (such as (CA)n or (CCTTAA)n).
They are found in a wide variety of eukaryotes and also in the chloroplastic genome of plants (Jarne
& Lagoda, 1996). Microsatellite
loci are non-coding DNA-sequences. Their exact function is unknown. They are
not subjected to selection and are highly polymorhpic.
Pedigree analyses have shown that they are codominant
and inherited in a Mendelian fashion. An individual
inherits alleles from both parents. All together, microsatellites
can be used to answer questions about genetic diversity within and between
populations, individual identification, distribution and numbers as well as
questions on population structure kinship structure, population dynamics and
taxonomy (Parker et al. 1998). In order to be able to do microsatellite
analyses, species specific primers are necessary. These primers attach to the
DNA surrounding the microsatellites and form new
strands of microsatellite DNA. Thus, enough DNA can
be synthesized to be able to visualize and analyze it.
Once amplification is done, the parts of
DNA with different molecular weights have to be separated from each other. This
is done by electrophoresis. The DNA is inserted in a gel through which a
current is passed. The DNA fragments with a low molecular weight will move
faster through the gel than the DNA fragments with a high molecular weight. By
adding a size standard, the molecular weights of the samples can be determined
and analyses can be done.
|
Protocol for collecting fecal samples in the field
used by the Primate Laboratory of the field station Los Tuxtlas
of UNAM At Los Tuxtlas we developed, after many
hours of field trials, a method for collecting and storing fecal samples for
DNA. Below is a description of material and method used is given. We suggest
that people rehearse collecting material before launching a field program. Material:
Sterile vials ( +/- 100 ml), Dehumidifier powder/cristals, Filter paper, Examination gloves, Sterile
plastic spoons, Colored flags, Permanent marker, Alcohol
Preparation:
Before going to the field, preparation of the vials has to be done. We use
100 ml sterile vials. Other vials can be used as well, but if they are not
sterile they have to be cleaned with alcohol. To each vial, approximately one
centimeter of a dehumidifier powder (e.g. silica gel or euqivalent)
is added. The dehumidifier dries the sample very quickly, and stops
degradation of the sample. The filter paper is cut in pieces of 10x10 cm,
folded and placed in the vial. All this work is done with gloves to reduce
contamination with human cells.
Fieldwork: Once we encounter a troop of howler monkeys in the field, we
determine sex and age of the animals present in the troop. Since howlers tend
to defecate at the same time, attention has to be paid at the locations of
the individual monkeys. Once a monkey defecates, we locate the fecal material
and place colored flags beside the feces. This way, we are able to
distinguish between fecal samples of different individuals, even when two
individuals defecated at the same spot. For each individual we use four vials
to collect fecal material in (for backup and use in other studies). In order
to work as sterile as possible, we use gloves to collect the samples. With a
sterile plastic spoon a small piece of fecal material (appr.
1x0,5x0,5 cm) is placed on the filter paper. The paper is then folded two
times, and the ends are folded into each other to make sure the dehumidifier
stays separated from the sample and no contamination can occur. Date, site
and individual are written on the lid of the vials and the four vials are
taped together and placed in a bag.
Once back from the field, the data of the samples we
collect are added in a database. Each vial is given a unique number and they
stored at room temperature for further analyses. |
|

Literature cited in tables above
- Marchant, L.F. et al, Highly successful non-invasive
collection of DNA from wild Chimpanzees.
- Escobar, P. (2000) Microsatellite
primers for the wild brown capuchin monkey Cebus
apella. Molecular Ecology 9, 107
- Zhang, X. et al. Extraction of DNA and PCR
analysis of DNA from free-ranging howling monkey (Alouatta palliata) feces.
- Mowat, G. et al. Using genetic tagging to estimate
animal population parameters (1999)
- Warner J.P.(1998) In
Molecular Genetic Analysis of Populations, a practical approach (ed. Hoelzel A.R.), pp 33. IRL Press,
Other related literature DNA fecal samples
Constable J.J, Packer
C., Collins D.A., Pusey A.E. (1995) Nuclear DNA from
primate dung. Nature 373-393
Wasser S., Houston C., Koehler G., Cadd
G., Fain S. (1997) Techniques for application of fecal DNA methods to field
studies of ursids. Molecular Ecology 6: 1091-1097
Kohn M.H. and Wayne R.K. (1997) Facts from feces
revisited. Tree 12: 223-227
Taberlet P., Waits L.P. and Lutkart
G. (1999) Noninvasive genetic sampling: look before you leap. Tree 14: 323-327
Taberlet P., Griffin S., Goossens
B., Questiau S., Manceau
V., Escaravage N., Waits L.P., Bouvet
J. (1996) Reliable geotyping of samples with very low
DNA quantities using PCR. Nucl. Acids Res. 24: 3189-3194
Frantzen M.A., Silk J.B., Ferguson J.W., Wayne
R.K., Kohn M.H. (1998) Empirical evaluation of preservation methods for fecal
DNA. Molecular Ecology 7 (10): 1423-8.
Abstract
Some other relevant references:
Beier P., Noss R.F. 1998. Do habitat corridors provide connectivity? Conservation Biology 12:6, 1241-1252
Bradley, BJ, Chambers, KE & Vigilant, L. 2001. Accurate DNA-based sex identification of apes using non-invasive
samples. Conservation Genetics
2: 179.181.
Brandley, BJ, Boesch, C & Vigilant, L. 2001. Identification and redesign of
human microsatellite markers for genotyping wild
chimpanzee (Pan troglodytes verus)
and gorilla (Gorilla gorilla gorilla)
DNA from feces. Conservation Genetics
1: 289-292.
Estrada A.E., Rivera A., Coates-Estrada R. 2002. Predation of artificial nests in a fragmented landscape in the
tropical region of Las
Frankham R. 1998. Inbreeding and extinction:
island populations. Conservation
Biology 12:3, 665-675
Morin PA, Chambers KE, Boesch,
C, Vigilant, L. 2001. Quantitative
polymerase chain reaction analysis of DNA from noninvasive samples for accurate
microsatellite genotyping of wild chimpanzees (Pan troglodytes verus). Molecular Ecology 10: 1835-1844.
Bradley, BJ & Viginlant,
L. 2002. False
alleles derived from microbial DNA pose a potential source of error in microsatellite genotyping of DNA from faeces.
Molecular Ecology News 2: 602-605.
Vigilant, L. Hofreiter, M, Siedel, H, Boesch, C. 2001. Paternity and relatedness in wild chimpazee communities. PNAS 96: 12890-12895.
Recent references for Alouatta:
James RA; Leberg PL; Quattro JM; Vrijenhoek RC. Genetic diversity in black howler monkeys (Alouatta
pigra) from
James RA;
Ellsworth JA; Hoelzer GA. Characterization of microsatellite loci in a
Torres OM; Leibovici M. [Characterization of
the karyotype of the red howler monkey Alouatta seniculus that inhabits
Bonvicino CR; Lemos B; Seuanez
HN. Molecular phylogenetics of
howler monkeys (Alouatta, Platyrrhini):
A comparison with karyotypic data..
CHROMOSOMA. 2001.
110(3), Pgs: 241-246
Torres OM; Leibovici M. [Characterization of
the karyotype of the red howler monkey Alouatta seniculus that inhabits
De Oliveira EHC; Suemitsu
E; Da Silva AF; Sbalqueiro IJ; Ando S; Someya K; Suzaki Y; Honda M. Geographical variation of chromosomal number in Alouatta fusca clamitans (Primates, Atelidae). CARYOLOGIA. 2000.
53(2), Pgs: 163-168
Reproductive
success increases with degree of kinship in cooperative coalitions of female
red howler monkeys (Alouatta seniculus).. BEHAVIORAL ECOLOGY AND SOCIOBIOLOGY.
2000. 48(4), Pgs:
253-267
de Oliveira EHC; de Lima MMC; Sbalqueiro
IJ; Pissinatti A. The karyotype
of Alouatta fusca clamitans from
Figueiredo WB;
Mudry MD; Rahn
M; Gorostiaga M; Hick A; Merani
MS; Solari AJRevised karyotype of Alouatta caraya (Primates: Platyrrhini)
based on synaptonemal complex and banding analyses. HEREDITAS. 1998.
128(1), Pgs: 9-16
Pope TR Effects of demographic change on group kin structure and gene dynamics
of populations of red howling monkeys. JOURNAL OF MAMMALOGY. 1998.
79(3), Pgs: 692-712
Consigliere S; Stanyon R; Koehler U; Agoramoorthy
G; Wienberg J Chromosome painting defines genomic
rearrangements between red howler monkey subspecies. CHROMOSOME
RESEARCH. 1996.
4(4), Pgs: 264-270
de Oliveira E H C; de Lima M M
C; Sbalqueiro IJ Chromosomal variation in Alouatta fusca. NEOTROPICAL PRIMATES. 1995.
3(4), Pgs: 181-183
Cortes-Ortiz L; Bermingham E; Rico C;
Rodriguez-Luna E; Sampaio I; Ruiz-Garcia M. Molecular
systematics and biogeography of the Neotropical monkey genus, Alouatta. MOLECULAR PHYLOGENETICS AND EVOLUTION. 2003. 26(1), Pgs: 64-81
de Oliveira EHC; de Lima MMC; Sbalqueiro
IJ; da Silva ASF Analysis of polymorphic NORs in Alouatta species (Primates,
Atelidae). CARYOLOGIA. 1999. 52(3-4), Pgs: 169-175
de Oliveria E H C Cytogenetic and phylogenetic
studies of Alouatta from Brasil
and
Rahn MI; Mudry M; Merani
MS; Solari AJ Meiotic behavior of the X1X2Y1Y2 quadrivalent of the primate Alouatta
caraya.CHROMOSOME RESEARCH. 1996. 4(5), Pgs: 350-356
de Oliveira EHC Cytogenetic
studies in the family Alouattinae. NEOTROPICAL
PRIMATES. 1995.
3(2), Pgs: 52-53
Mudry MD; Rahn IM; Solari
AJ Meiosis and chromosome painting of sex chromosome systems in Ceboidea. AMERICAN
JOURNAL OF PRIMATOLOGY. 2001.
54(2), Pgs: 65-78
Schneider
H; Canavez FC; Sampaio I; Moreira MAM; Tagliaro CH; Seuanez HN Can molecular data place each neotropical monkey in its own branch? CHROMOSOMA.
2001. 109(8), Pgs:
515-523
Stanyon R; Consigliere S; Bigoni F;
Ferguson-Smith M; O'Brien PCM; Wienberg J Reciprocal
chromosome painting between a New World primate, the woolly monkey, and humans.
CHROMOSOME RESEARCH. 2001. 9(2), Pgs: 97-106
Escobar-Paramo P Microsatellite primers
for the wild brown capuchin monkey Cebus apella. MOLECULAR ECOLOGY. 2000. 9(1), Pgs: 107-108
Groves
C Why taxonomic stability is a bad idea, or why are
there so few species of primates (or are there?). EVOLUTIONARY
ANTHROPOLOGY. 2001.
10(6), Pgs: 192-198
Romagno D Primate
tables chromosome. CARYOLOGIA. 2001. 54(4), Pgs: 285-297
Dietz
JM Conservation of biodiversity in neotropical
primates. BIODIVERSITY, II. UNDERSTANDING AND PROTECTING OUR
BIOLOGICAL RESOURCES. M.L. Reaka-Kudla, D.E.
Wilson, E.O. Wilson, Editors.
Pope TR Socioecology, population
fragmentation, and patterns of genetic loss in endangered primates.
CONSERVATION GENETICS: CASE HISTORIES FROM NATURE. J.C. Avise,
J.L. Hamrick, Editors.
Pope TR Socioecology, population
fragmentation, and patterns of genetic loss in endangered primates.
CONSERVATION GENETICS: CASE HISTORIES FROM NATURE. J.C. Avise,
J.L. Hamrick, Editors.
Gibbons
E F Jr Conservation of primates in captivity. CONSERVATION OF ENDANGERED SPECIES IN CAPTIVITY. E.F.
Gibbons, Jr., B.S. Durrant, A.J. Demarest, Editors.
|
Contribuye a la conservación de las selvas tropicales |
Protege los primates Mesoamericanos |
copyright@alejandro estrada